9月1日,9570金沙登录入口白明义教授课题组在Nature Plants发表了题为Regulatory functions of cellular energy sensor SnRK1 for nitrate signalling through NLP7 repression的研究论文。该研究揭示了SnRK1感受植物体内碳水化合物和硝态氮的代谢变化,调控硝态氮信号转导,进而维持植物碳-氮平衡的分子机理。
氮是植物需求量最大的矿质元素,在作物生产中起着决定生物量和产量的关键作用。植物根系中氮的吸收利用效率受到地上部叶片光合作用碳同化能力的显著影响,二者相互依赖以维持植物碳-氮平衡,来保证植物适应多变的生存环境,但植物调控碳-氮平衡的分子机理还不清楚。
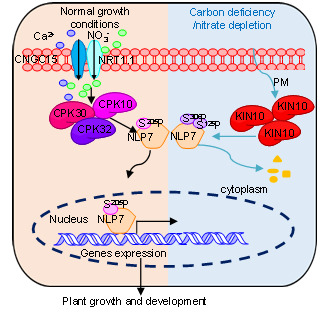
植物的SNF1-Related Kinase1 (SnRK1)是一种进化上保守的能量感受蛋白激酶,当能量供应有限时,可以协调转录调控网络以维持细胞的能量稳态。白明义课题组前期的研究工作显示SnRK1能感受植物体内碳水化合物的代谢变化调控植物气孔发育来增强植物的环境适应性。为进一步解析SnRK1调控植物发育和环境适应性的分子机理,白明义课题组利用酵母双杂交筛选了SnRK1催化亚基KIN10的相互作用蛋白,其中发现植物硝态氮信号转导的关键转录因子NLP7是KIN10的相互作用蛋白。体内和体外的实验证实KIN10与NLP7相互作用。硝态氮作为信号分子能快速诱导下游基因的表达,但光照强度降低或光照时间缩短引起的光合产物匮乏会抑制硝态氮对下游基因的表达调控,但这种抑制效果在kin10突变体中被显著减弱。过表达KIN10抑制了植物对硝态氮的响应,使得大约34.5%的氮响应基因不再被硝态氮调控。这些结果表明,KIN10是植物感知光合产物匮乏调控硝态氮响应的关键组分,是联系光合作用与硝态氮信号的重要枢纽。
NLP7是植物氮响应的关键转录因子。硝态氮处理会诱导NLP7转移到细胞核,但在光合产物匮乏时,硝态氮诱导NLP7进核的能力显著降低。KIN10过表达会导致NLP7滞留在细胞质不能进核行使功能。同时,过表达KIN10还会促进NLP7蛋白降解。KIN10磷酸化NLP7的第125位和第306位的丝氨酸。NLP7S125AS306A(模拟不能被KIN10磷酸化的形式)能恢复nlp7突变体氮敏感的表型,而NLP7S125DS306D(模拟被KIN10磷酸化的形式)则不能恢复nlp7突变体的表型。硝态氮处理会诱导NLP7S125AS306A定位在细胞核调控氮响应基因表达,而NLP7S125DS306D主要定位在细胞质,不能进核发挥功能。此外,在光合作用产物匮乏时会诱导NLP7蛋白发生降解,但对NLP7S125AS306A蛋白没有显著影响。以上结果表明,KIN10磷酸化NLP7使其滞留在细胞质降解,进而抑制植物硝态氮信号转导。
值得注意的是,白明义课题组先前的研究工作显示植物激素油菜素甾醇(Brassinosteroid,BR)信号转导关键转录因子HBI1通过调控活性氧的稳态促进硝态氮信号转导,并证明活性氧处理会导致NLP7滞留在细胞质不能进入细胞核来行使功能,但活性氧调控NLP7亚细胞定位的分子机制并不清楚。最近该课题组在Nature Communications发表工作证明活性氧通过抑制KIN10与KINβ的相互作用,促进KIN10转运到细胞核来行使功能。结合该研究,研究人员推测逆境胁迫条件下积累的活性氧会促使KIN10进入到细胞核,核中积累的KIN10磷酸化NLP7,导致NLP7转运到细胞质然后降解,从而抑制植物硝态氮信号转导。此外,该课题组还有工作显示BR信号转导中的负调控激酶BIN2能磷酸化KINβ,减弱KIN10与KINβ的相互作用促进KIN10进入细胞核抑制氮信号,而BR通过抑制BIN2的活性使得KIN10滞留细胞质,从而促进氮信号。
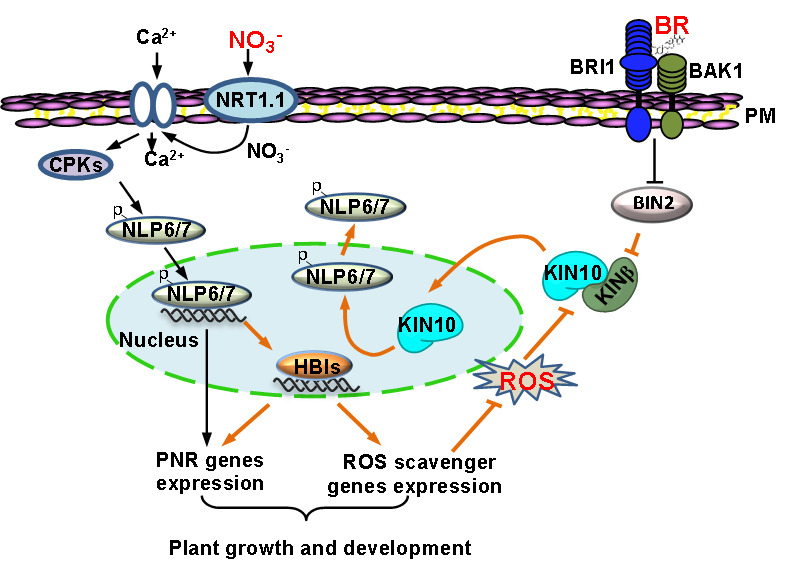
综上所述,SnRK1激酶作为植物体内中枢能量感受器,不仅能被碳水化合物和氮代谢所调控,还能被活性氧所激活,而被植物生长促进激素BR所抑制。SnRK1整合植物体内的代谢信号、内源激素以及外源环境刺激来调控NLP7活性,进而调控植物硝态氮信号。
9570金沙登录入口的博士研究生王红蕾,副教授韩超为该论文的共同第一作者,白明义教授为该论文的通讯作者。山西农业大学的王家刚教授,褚晓茜副教授,浙江省农业科学研究院的邓志平研究员参与了该项研究工作。该研究工作得到国家自然科学基金,山东省良种工程,山东省重大基础研究项目、9570金沙登录入口青年交叉创新群体项目和9570金沙登录入口未来青年学者项目的支持。
